УДК 577.71:519.95
Старение и продолжительность жизни: от мушек до человека.
Институт проблем управления РАН, e-mail: novoselc@ipu.rssi.ru
Одни и те же демографические процессы происходят в настоящее время во всех развитых странах мира - продолжительность жизни увеличивается, количество пожилых людей возрастает, а число рождающихся детей падает [1]. Считается, что Европа уже вступила в эпоху негативных популяционных изменений - тенденция к изменению возрастного состава населения в сторону увеличения пожилых сохранится, даже если фертильность населения немедленно увеличится до уровня, необходимого для естественного воспроизводства популяции [2]. Такие тенденции вызывают обоснованное беспокойство, поскольку в связи с увеличением доли пожилых лиц во всех странах возникает опасность чрезмерного увеличения социальной нагрузки на молодую и работоспособную часть населения. Доля пожилых лиц в 15 странах Европейского Сообщества с 20 % в 2000 г. вырастет в течение ближайших 20 лет до 33-34 % [2]. Кроме того, из-за общего постарения населения возрастают риски заболеваемости и инвалидизации.
Начиная с 1840-х гг. максимальная продолжительность жизни в мире увеличивалась по линейному закону со средней скоростью 2.5 года за десятилетие (рис. 1). Различные страны поочередно играют роль лидеров, возглавляя гонку. То, что страны, ранее не входившие в число ведущих, со временем вырываются вперед, означает, что продолжительность жизни в них во время "рывка" увеличивается значительно быстрее, чем у страны-лидера [4]. В настоящее время нет видимых причин, по которым этот рост мог бы прекратиться.
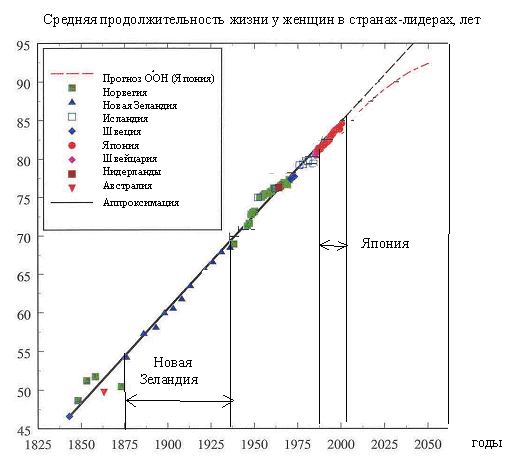
Рис. 1. Изменение средней продолжительности жизни женщин в странах-лидерах ([3], с изменениями). За сто шестьдесят лет продолжительность жизни увеличилась на 40 лет, причем постоянно происходила смена страны-лидера. С 1875 и до Второй Мировой войны лидировала Новая Зеландия, а в последнее время вперед вырвалась Япония, в которой средняя продолжительность жизни женщин превысила 85 лет.
В последние годы все чаще разгораются дискуссии о том, ограничены ли вообще пределы жизни. В этих дискуссиях высказываются прямо противоположные точки зрения. Дж. Вопель утверждает, что рост средней продолжительности жизни, показанный на рис. 1, будет продолжаться и далее [3], а его главный оппонент С. Дж. Ольшански говорит, что средняя продолжительность жизни уже достигла своего предела и дальнейший ее рост невозможен [5].
В связи с тем, что данные по старению человека довольно ограничены, в этих дискуссиях большую роль играют результаты, полученные на животных. Так, американский биодемограф Дж. Кэри, проведя опыты с популяцией средиземноморских плодовых мушек Medfly объемом в миллион особей, обнаружил удивительные факты. Оказалось, что при средней продолжительности жизни мушек около 50 дней самая долгоживущая мушка прожила более 200 дней [6]. Мало того, было обнаружено, что смертность (т.е. вероятность умереть в течение ближайшего отрезка времени) у всех живых существ с увеличением возраста выше некоторого предела сначала начинает замедляться, а затем даже падать [7]. Хотя эти данные пока плохо распространяются на человека по причине недостаточной величины имеющейся выборки, имеются веские основания для того, чтобы в этом не сомневаться. Как эти, так и другие подобные результаты привели Дж. Кэри к мысли об отсутствии естественного предела продолжительности жизни в природе [8].
Мы тоже считаем, что человечество еще далеко не достигло максимума средней продолжительности жизни. В принципе, если человек не болеет и не попадает в ситуации, связанные с риском преждевременной смерти, то максимальная продолжительность его жизни должна составлять около 140 лет [9]. В 2004 г. журнал американский журнал "Journal of Gerontology" провел дискуссию, посвященную вопросам старения и продолжительности жизни. В дискуссии приняли участие ведущие специалисты из разных стран мира. Так, известный американский биолог Леонард Хейфлик утверждает, что никакие вмешательства не могут замедлить, остановить или обратить процесс старения у человека [10]. Что касается увеличения продолжительности жизни, говорит он, то она происходит исключительно за счет сокращения смертности от детских и старческих болезней. Механизм старения - общий для всех живущих на Земле существ - остается пока не затронутым и в настоящее время вообще не ясно, можно ли на него каким-либо образом воздействовать.
Таким образом, критическое постарение населения в Европе и других странах мира поставило в повестку дня вопрос о механизмах старения. Существует ли связь между процессами увеличения продолжительности жизни и сокращения деторождения? В биодемографической литературе эта проблема анализируется с точки зрения распределения ресурсов организма между двумя процессами: поддержания организма и производства потомства, которые рассматриваются в качестве конкурентов. Каковы естественные механизмы, управляющие процессом "переброса" и использования энергии, освобождающейся у организмов, которые не рождают потомков? Приводит ли само по себе сокращение рождаемости к увеличению продолжительности жизни? Для животных этот факт многократно доказан экспериментально, но для человека пока имеются лишь отдельные публикации, подтверждающие его [11]. До сих пор не ясен вопрос о том, одинаковы ли механизмы, приводящие к увеличению продолжительности жизни у человека и у экспериментальных животных, которые находятся в "идеальных" условиях жизни.
Эти положения говорят о том, что деторождение тесно связано с увеличением продолжительности жизни. В этой связи большую тревогу у демографов вызывает сокращение деторождения. Как в Европе, так и в США, Японии и Канаде количество детей, рождаемых в среднем каждой женщиной на протяжении ее жизни, в настоящее время составляет примерно 1,4. В то же время известно, что для сохранения постоянства уровня населения (без учета миграции) необходимо, чтобы каждая женщина рожала чуть больше 2 детей. Именно поэтому резкое изменение структуры населения ожидается в ближайшие десятилетия в крупнейших регионах (Европы и, возможно, Америки). Континенты Африка и Азия уйдут далеко вперед, обогнав нынешние центры цивилизации. Иммиграция в любых мыслимых пределах не в силах предотвратить эту тенденцию. Сегодня демографы боятся последствий этого поворота событий и рискуют прогнозировать последствия лишь при условии, что деторождение в Европе немедленно возрастет до 1,5 -1,8 [2]. Даже при таких оптимистических предположениях население пятнадцати ведущих стран Европы к 2050 г. уменьшится с нынешних 375 млн. до 300 млн. человек [10].
Существующие в Европе представления об идеальном размере семьи никак не отражают этих реалий и не учитывают угроз [12].
Конечно, представленные данные носят описательный характер. Более глубинные процессы, связанные с причинами этих явлений и с возможными способами воздействия на них, до сих пор находятся в зачаточном состоянии. Хотя в последнее время общественный интерес к проблемам демографии и к наукам, посвященным анализу процессов старения (геронтологии и биогеронтологии) и медицине для пожилых людей (гериатрия) резко возрос, механизмы управления этими процессами до сих пор неясны. В настоящее время специалисты различных дисциплин во всем мире стараются понять, как связаны причины этих изменений с физиологическими и генетическим факторами. Расшифровка генома человека и дрозофилы показала удивительную их близость. Около 80 % генов оказались общими, что подтверждает важность экспериментов на животных и возможность использования получаемых данных для человека [13]. Число публикаций, в которых анализируются проблемы человеческих болезней на материале мушиных популяций, с начала XXI в. резко увеличилось (рис. 2).
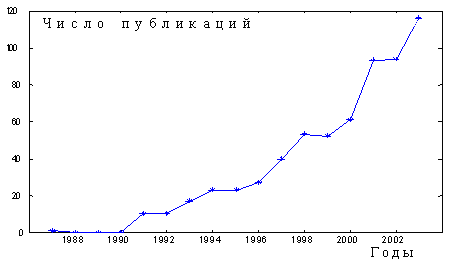
Рис.2. Рост числа ежегодных публикаций в ведущих журналах мира, посвященных исследованию старения и развития заболеваний у человека на основе анализа популяций плодовой мушки.
Проблемы старения и долгожительства перестают быть достоянием только демографов, биогеронтологов и гериатров. Они становятся предметом анализа все более широкого круга наук, в том числе теории управления [14, 15]. Предметом анализа наших работ явилось исследование индивидуальной плодовитости различных плодовых мушек - в частности, дрозофилы D. melanogaster и средиземноморской плодовой мушки Medfly [16].
Обычно в исследуемых популяциях мушек плодовитость популяции выглядит так, как показано на рис. 3. Сначала число яиц, сносимых всеми мушками в популяции, возрастает, а затем начинает убывать.
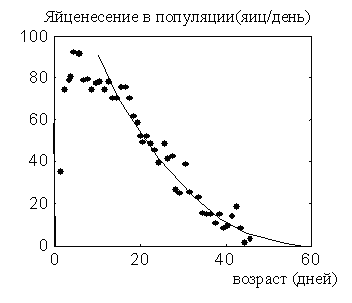
Рис. 3. Среднепопуляционная плодовитость популяций дрозофилы [17]. Показана экспонента, которой аппроксимируются график яйценесения в зрелых возрастах.
Однако более тщательный анализ экспериментальных данных показывает, что это не так [14, 15]. Взрослая мушка начинает свой жизненный путь со взросления (в это время она не несет яиц, накапливая их в овариолах). Начав нести яйца в возрасте x =Xonset, она несет их с постоянной скоростью вплоть до исчерпания жизненного ресурса. Этот период мы называем периодом взрослости. Когда энергетические ресурсы начинают иссякать, начинается старость - темп яйценесения начинает экспоненциально снижаться (рис. 4).
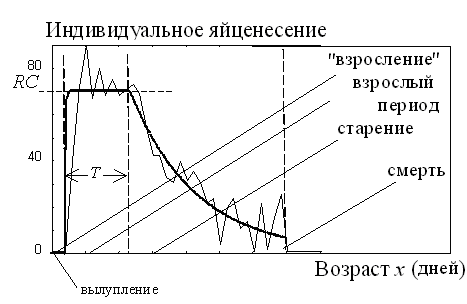

Тогда график среднепопуляционного яйценесения, показанный на рис. 3, получается путем усреднения графиков рис. 4. На рис. 5 показано, как такое усреднение происходит.
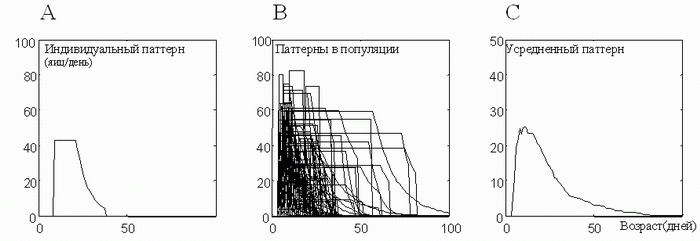
Рис. 5. Усреднение плоских индивидуальных графиков (А) дает картину, наблюдающуюся в популяциях (С). Экспоненциальный "хвост" индивидуального паттерна становится более длинным, а короткие периоды зрелости, усредняясь, дают максимум среднепопуляционного яйценесения. Таким образом, этот максимум является артифактом усреднения [14].
Анализируя процессы яйценесения у мушек дрозофилы и Mеdfly, мы отвечаем на следующие вопросы [16]:
- существует ли точка начала старения в популяциях?
- действительно ли график индивидуального яйценесения является плоским?
- действительно ли яйценесение в старости затухает по экспоненте?
Чтобы получить ответы, мы аппроксимируем все индивидуальные паттерны яйценесения в популяции Wayne State методом наименьших квадратов и затем усредняем их по всем мухам. Для того, чтобы проверить наличие в популяции точки начала старения, мы "центрируем" все индивидуальные паттерны. Для этого мы располагаем каждый график так, чтобы предсказанная методом наименьших квадратов точка начала старения всегда попадала в "начало координат". Обозначим символом r(i, j) число яиц, откладываемых мушкой j в день i. Усредненная разность Di между реальный числом снесенных яиц и соответствующим уровнем плато RC имеет вид
(1) | 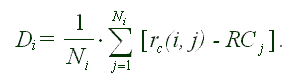 |
Здесь Di - усредненная "центрированная" разность для возраста i, а величина rc(i, j) = r(Xonset,j - Tmax + Tj + i, j) представляет собой "центрированный" отсчет яйценесения мушки j в день i.
График Di для популяции дрозофил Wayne State [16] дан на рис. 6 сверху, а число мушек Ni, по которому производится усреднение, приведен на рис. 6 внизу. Усредняясь по 493 мушкам, итоговый паттерн принимает характерный двухчастный вид. Левее центральной точки все ошибки усреднились, давая нулевой уровень, а вправо от этой точки усреднение дает характерную кривую, близкую к экспоненте. График для популяции мушек Medfly выглядит аналогично (график не показан). Таким образом, возраст начала старения, делящий паттерн яйценесения на две части, действительно существует.
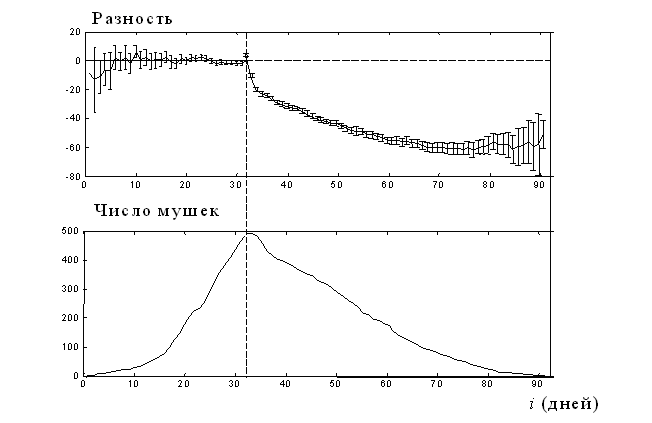
Рис. 6. График, демонстрирующий существование начала старения у дрозофил.[16]. Вверху: усредненные разности для популяции дрозофил Wayne State. Вертикальный пунктир показывает положение центральной точки, полученной у каждого индивидуума методом наименьших квадратов. Все усредняемые паттерны центрированы к дню 32. Фактически Tmax = 34 дней, но чтобы сделать результат чище, два первых дня из каждого паттерна перед усреднением удалены. Показаны 95 % доверительные интервалы. Внизу: число мушек, по которым проводилось усреднение в каждом из возрастов.
Покажем теперь, что в периоде взрослости паттерн имеет плоский вид. Чтобы получить ответ на этот вопрос, мы усредним индивидуальные ошибки между случайными отсчетами у каждой мушки и ее собственным уровнем плато. Это позволит нам оценить степень близости регулярных паттернов и их исходных прототипов в среднем.
Поскольку в каждый день взрослости мы усредняем только ошибки для взрослых индивидуумов, мы получаем
(2) | 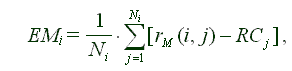 |
где EMi - ошибка на i-тый день взрослости (Error of Maturity), rM(i, j) = r(Xonset,j+ i, j), а (rij RCj) представляет собой индивидуальную ошибку у мушки j в этот день. Тогда j = 1, ... , Ni, где Ni - число взрослых мушек в день i [18].
Ошибки EMi для 493 мушек Drosophila и 1170 средиземноморских плодовых мушек показаны на рис. 7 (сверху). Действительно, на графиках отсутствует максимум. У мушек Drosophila максимальное отклонение от среднего составляет +7 яиц/день и достигается на 4 день. Эта ошибка составляет 11.5 % среднего уровня, равного 60.47 яиц/день. Аналогичные цифра справедливы и в другом случае. Доверительные интервалы к концу плато увеличиваются из-за того, что число мушек уменьшается.
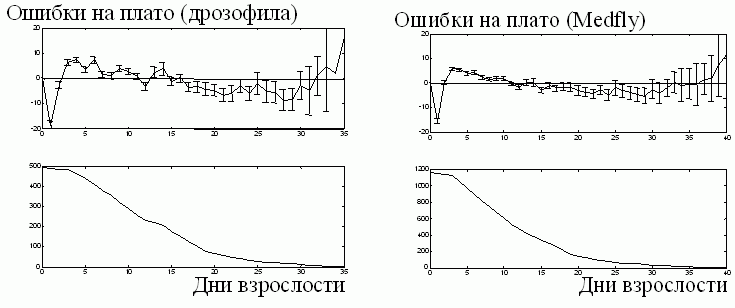
Рис. 7. Систематические ошибки на плато в популяции Wayne State Drosophila [18] и у 1170 средиземноморских плодовых мушек [16]. Все точки начала репродукции сдвинуты в начало координат. Сверху: усредненные ошибки на первый, второй и т.д. день периода взрослости по сравнению с соответствующим уровнем плато. Внизу число взрослых мушек. Показаны 95 % доверительные интервалы.
Оба графика практически плоские. Особенности их структуры рассмотрены в другом месте (Novoseltsev et al, в печати). Таким образом, мы подтвердили вывод о наличии плато в индивидуальном яйценесении плодовых мушек.
Чтобы ответить на третий вопрос, надо вычислить индивидуальные ошибки на периоде старости и сравнить их с соответствующими величинами убывающего "хвоста" яйценесения. После этого ошибки надо усреднить. Мы имеем
(3) | 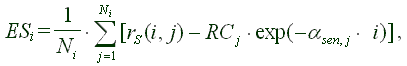 |
где ESi - ошибка на i-й день старения (Error of Senescence), rS(i,j)=r(Xsen,j + i, j), Ni представляет собой число стареющих мух в день i, а αjsen - темп старения мушки с номером j. Остальные обозначения те же, что и в уравнении (2).
Эти ошибки показаны для популяции дрозофил на рис. 8 (сверху). Все точки начала старения перемещены в начало координат. Для средиземноморских мушек графики выглядят аналогично (не представлены). Таким образом, мы положительно отвечаем на вопрос об экспоненциальном характере снижения яйценесения в период старости у мушек [16].
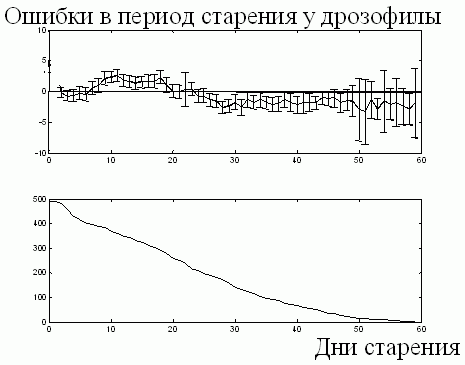
Рис. 8. Систематические ошибки у дрозофилы в период старения (сверху) и число мушек, по которым производится усреднение (снизу). Обозначения те же, что и выше.
Анализ изменения параметров индивидуального яйценесения позволяет сделать далеко идущие выводы относительно аналогии механизмов управления плодовитостью у дрозофилы и биологических механизмов ограничения рождаемости у человека. Оказывается, что период взрослости, т.е. постоянства темпа яйценесения у дрозофил (в отличие, например, от средиземноморской плодовой мушки) имеет тенденцию к постоянству. Этим он напоминает генетические ограничения максимального возраста деторождения у женщин.
Рассмотрим эти аналогии несколько более подробно. На рис. 4 показан типичный паттерн индивидуального яйценесения у дрозофилы. Среди параметров, его описывающих, выделим три основных - интенсивность яйценесения (RC, число яиц в день) на периоде взрослости, продолжительность этого периода (T) и показатель экспоненты снижения темпа яйценесения в старости ( αsen).
Мы уже говорили о том, что индивидуальное яйценесение у мушек сильно флуктуирует. Поэтому для улучшения качества дальнейшего анализа мы сгруппируем отдельных индивидуумов дрозофилы в децильные кластеры по продолжительности их жизни. Возьмем теперь средние по кластерам значения основных параметров (RC, T и αsen) и изобразим их на графиках (см. рис. 9). Оказывается, что два их этих параметров (RC и T) практически не меняются, а третий (αsen) меняется очень сильно.
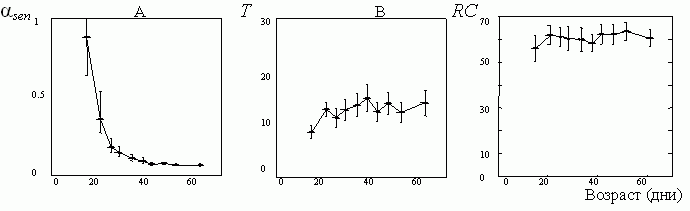
Рис. 9. Среднекластерные значения индивидуальных параметров, описывающих яйценесение у дрозофилы [16]. Оказывается, что только αsen сильно меняется между кластерами (А), тогда как два других (RC [яиц/день]и T[дней]) от кластера к кластеру практически не меняются (В и С).
Из этого следует, что у дрозофилы действительно существует тенденция к сохранению постоянства периода взрослости T. Правда, небольшой объем выборки (несколько менее 500 индивидуумов) делает этот вывод предварительным. Чтобы сделать его более наглядным и убедительным, мы в каждом из кластеров подобрали таких индивидуумов, у которых продолжительность периода взрослости практически совпадает со среднекластерными значениями. Соответствующие паттерны яйценесения представлены на рис. 10.
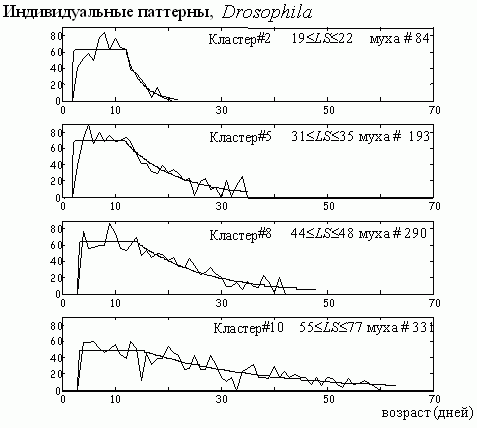
Рис. 10. Индивидуальные паттерны яйценесения у дрозофилы [16]. Показаны представители кластеров 2, 5, 8 и 10, имеющие примерно одинаковый параметр T и приведены значения продолжительности жизни LS в этих кластерах.
В любом случае, если этот вывод будет подтвержден дальнейшими исследованиями, он окажется весьма важным. Тогда вопрос об изучении развития различных заболеваний у человека с помощью исследований на дрозофиле [13] можно будет дополнить изучением проблемы деторождения, что представляется особенно важным в свете тех демографических тенденций, о которых говорилось в начале настоящей статьи.
Литература
1. Lutz W., O'Neill B.C., Schеrbov S. Europe's population at a turning point // Science. -2003.- 299:1991-1992.
2. Goldstein J., Lutz W., Scherbov S. Long-term population decline in Europe: the relative importance of tempo effects and generation length // Pop. Dev. Review. -2003.-29: 699-707.
3. Oeppen J., Vaupel J.W. Demography. Broken limits to life expectancy // Science. -2002. -296: 1029-1031.
4. Arking R., Novoseltsev V., Novoseltseva J. The human life span is not that limited: the effect of multiple longevity phenotypes // Journ. Geront. Biol. Ser. -2004. -59a: 697-704.
5. Carnes, B.A., Olshansky S.J., Grahn D. Biological evidence for limits to the duration of life // Biogerontology. -2003. - 4:31-45.
6. Carey JR. How Mediterranean fruit flies resist aging, live long and remain fertile. In: Paradoxes of longevity / Robine J.-M., Forette B., Franceschi C. and Allard M., Eds.-N.Y., 1999.- P. 23-34.
7. Vaupel JW., Carey J.R., Christensen K., Johnson T.E.,Yashin A.I., Holm N.V., Iachine I.A., Khazaeli A.A., Liedo P., Longo V.D., Zeng Y., Manton K.G., and Curtsinger J.W. Biodemographic Trajectories of Longevity// Science. - 1998. - 280: 855-860.
8. Carey J.R. Insect biodemography // Ann. Rev. Entomol. -2001. - 46:79-110.
9. Новосельцев В.Н. Математическое моделирование организма // Наука в России. - 2003. -1:52-58.
10. Hayflick L. "Anti-aging" is an oxymoron // Journ. Geront. Biol. Ser. - 2004. - 59a: 573-578.
11. Gavrilova N.S., Gavrilov L.A., Semyonova V.G., Evdokushkina G.N. Does exceptional human longevity come with a high cost of infertility? // Ann. N.Y. Acad. Sci. - 2004. - 1019: 513-517.
12. Goldstein J., Lutz W., Testa M.R. The emergence of sub-replacement family size ideals in Europe // Pop. Res. Policy Review. - 2003.- 22:479-496.
13. O'Kane C.J. Modelling human diseases in Drosophila and Caenorhabditis // Seminars in Cell & Dev. Biol. - 2003. - 14:3-10.
14. Novoseltsev V.N., J.A.Novoseltseva, A.I Yashin. What does a fly individual fecundity pattern look like? The dynamics of resource allocation in reproduction and ageing // Mechanisms of ageing and development. - 2003. - 124: 605-617.
15. Novoseltsev V.N., J.R. Carey, J.A. Novoseltseva, N.T. Papadopulos, S. Blay, A.I. Yashin. Systemic mechanisms of individual reproductive life history in female Medflies // Mechanisms of ageing and development. - 2004. - 125(1): 77-87.
16. Novoseltsev V.N., R. Arking, J. R. Carey, J.A. Novoseltseva, A.I. Yashin. Individual fecundity and senescence in Drosophila and Medfly. Presented to Journ. of Gerontol.
17. Partridge L., K.Fowler. Direct and correlated responses to selection on age at reproduction in Drosophila melanogaster // Evolution. - 1992. - 46: 76-91.
18. Novoseltsev V.N., R. Arking, J.R. Carey, J.A. Novoseltseva, A.I. Yashin. How an individual fecundity looks in Drosophila and Medflies // Ann. N.Y. Acad. Sci. - 2004. - 1019: 577-580.